|
||||
|
|
Глава XIVМолекулярная биология: нуклеиновая кислота Вирусы и геныИтак, молекула белка стала управляемой. И вдруг совершенно неожиданное, поразительное открытие: химическая основа жизни вовсе не молекула белка, а другая частичка. Только когда принялись за изучение природы фильтрующихся вирусов, стала ясна огромная важность этого открытия. Природа вирусов представляла загадку для целого поколения. Известно, что вирусы вызывают заболевания, были даже разработаны методы борьбы сними. Однако физические свойства вирусов все еще оставались неизвестными. Решающую роль в определении размера вирусов сыграло изобретение фильтров достаточно мелкопористых, чтобы задерживать вирусные частицы. Вирусы оказались немного меньше, чем мельчайшие из известных клеток, но значительно больше самой крупной белковой молекулы. Разглядеть вирусы позволил лишь электронный микроскоп. Их размеры варьируют в широких пределах, начиная от вирусов — мельчайших точек — и до сравнительно крупных структур строго геометрической формы с различимым внутренним строением. К наиболее крупным вирусам относятся бактериофаги, которые «охотятся» за мелкими микроорганизмами; некоторые фаги имеют хвостики и напоминают крошечных головастиков. Крупнее вирусов, но мельче бактерий риккетсии, названные так в честь Риккетса. Риккетсии вызывают, в частности, пятнистую лихорадку Скалистых гор — заболевание, изученное бактериологами. Возник вопрос, являются ли вирусы живыми организмами. В 1935 г. американский биохимик Уэнделл Мередит Стенли (род. в 1904 г.), работая с экстрактом вируса табачной мозаики, получил игольчатые кристаллы. Оказалось, что эти кристаллы обладают высокой инфекционностью. Другими словами, ученый получил вирус в кристаллическом виде, а живые кристаллы — явление трудно объяснимое. С другой стороны, нельзя ли допустить, что клеточная теория неточна и что клетки не являются неделимыми единицами жизни? Вирус много мельче клетки и в противоположность ей ни при каких условиях не способен существовать независимо. Однако вирусу удается проникнуть в клетку, размножиться там и в некоторых основных проявлениях вести себя, как живое существо. Нет ли каких-либо внутриклеточных образований, каких-либо доклеточных элементов, которые были бы действительной основой жизни — структурой, управляющей остальной частью клетки? Не является ли вирус таким клеточным компонентом, когда-то и как-то отщепившимся от клетки, но готовым заселить ее и сделать чуждой истинному «хозяину»? Если это так, такие доклеточые компоненты должны были бы находиться и в нормальных клетках. Кандидатами на эту роль, вероятнее всего, следует считать хромосомы. В первые годы нашего столетия стало очевидным, что хромосомы несут в себе факторы, управляющие наследованием физических свойств. Это определяет их руководящее положение в клетке, как и можно было ожидать от ключевых доклеточных компонентов. Однако хромосома значительно крупнее вируса. Но число хромосом гораздо меньше количества наследуемых признаков. Отсюда можно было сделать вывод, что одна хромосома состоит из многих, возможно тысяч, частиц, каждая из которых управляет отдельным признаком. Эти отдельные частицы датский ботаник Вильгельм Людвиг Иогансен (1857–1927) в 1909 г. назвал генами (в переводе с греческого — дать жизнь чему-либо). Однако в первое десятилетие XX в. отдельного гена, как и отдельных вирусов, еще не удавалось увидеть, хотя его проявления довольно успешно наблюдались. Ключ к этим исследованиям подобрал американский генетик Томас Хант Морган (1866–1945), использовав в 1910 г. новый биологический объект — плодовую мушку дрозофилу. Это маленькое насекомое неприхотливо, довольно легко размножается; кроме того, наличие в клетках дрозофилы всего четырех пар хромосом облегчает исследования. Изучая эту мушку поколение за поколением, Морган обнаружил огромное количество мутаций. Ему удалось показать, что различные признаки связаны, то есть наследуются как один комплекс. Значит, гены, управляющие этими признаками, должны находиться на одной хромосоме, которая и наследуется как целое. Но сцепленные друг с другом признаки связаны не на век. Бывает, что один из признаков наследуется без связи с другим. Это происходит потому, что пары хромосом случайно обмениваются участками (кроссинговер), так что целостность отдельной хромосомы не абсолютна. Подобные опыты позволили определить место каждого конкретного гена на хромосоме. Чем больше расстояние между двумя генами, тем больше вероятность перекрещивания произвольно расположенных генов. Изучая частоту, с которой расщепляются два особым образом связанных признака, можно определить относительное положение генов. В 1911 г. была составлена первая карта расположения генов в хромосомах (для дрозофилы). Один из учеников Моргана, американский генетик Герман Иозеф Мёллер (1890–1967), предложил метод увеличения частоты мутаций (1919). Он обнаружил, что повышение температуры увеличивает частоту мутаций. Это не было результатом общего «перемешивания» генов. Всегда оказывалось, что поражался один ген, тогда как его дубль на другой хромосоме данной пары оставался нетронутым Мёллер пришел к выводу, что эти изменения происходят на молекулярном уровне. Следующим шагом в его исследованиях было применение рентгеновских лучей, обладавших более высокой энергией, чем легкое нагревание. Отдельный рентгеновский луч, попав в хромосому, действует на нее в определенной точке. И действительно, в 1927 г. Мёллеру удалось доказать, что рентгеновские лучи значительно повышают темп мутирования. Эти исследования продолжил американский ботаник Альберт Фрэнсис Блэксли (род. в 1874 г.). В 1937 г. он показал, что темп мутаций можно повысить, действуя специфическими веществами (мутагенными факторами). Лучшим мутагенным фактором оказался колхицин — алкалоид, выделенный из безвременника (семейство ирисовых). Таким образом, к середине 30-х годов и вирусы и гены утратили покров таинственности. И те и другие оказались молекулами примерно одной и той же величины и близкой химической природы. А нельзя ли гены считать «прирученными» клеточными вирусами? И может ли вирус быть «диким геном»? Роль ДНККак только получили кристаллическую форму вирусов, стало возможным вести исследования по методу дифракции рентгеновских лучей. Вирусы, безусловно, относились к белкам, будучи особой их разновидностью, носящей название нуклеопротеидов. Успехи техники окрашивания препаратов позволили выяснить химическую природу отдельных субклеточных структур. Было установлено, что хромосомы (а следовательно, гены) также относятся к нуклеопротеидам. Молекула нуклеопротеида состоит из молекулы белка, связанной с фосфорсодержащим веществом, известным под названием нуклеиновой кислоты. Впервые нуклеиновые кислоты открыл в 1868 г. швейцарский биохимик Фридрих Мишер (1844–1895) в ядрах клеток гноя. Долгое время их считали специфически ядерным компонентом. Когда оказалось, что нуклеиновые кислоты присутствуют и вне ядер, уже поздно было менять название. Нуклеиновые кислоты подробно изучил немецкий биохимик Альбрехт Коссель (1853–1927), которому в 1880 г. удалось расщепить их на более мелкие составные части, включавшие фосфорную кислоту и сахар, точного состава которых он не смог определить. Кроме того, в нуклеиновой кислоте он обнаружил два соединения класса пуринов, молекулы которых представляли циклические соединения с двумя кольцами, содержащими четыре атома азота. Эти вещества Коссель назвал аденином и гуанином (а иногда просто обозначал буквами А и Г). Он обнаружил также три пиримидина (вещества с одним кольцом, содержащие два атома азота), которые были названы им цитозином, тимином и урацилом (Ц, Т и У). Американский химик Фебус Арон Теодор Левин (1869–1940), изучая эти вещества на протяжении 20-х и 30-х годов, показал, что в молекуле нуклеиновой кислоты молекула фосфорной кислоты, молекула сахара и молекула одного из пуринов или пиримидинов образуют трехкомпонентное соединение, которое он назвал нуклеотидом. Молекула нуклеиновой кислоты состоит из цепочки этих нуклеотидов, подобно тому как молекула белка — из цепей аминокислот. Нуклеотидная цепь построена так, что молекула фосфорной кислоты одного нуклеотида связана с сахарной группировкой соседнего нуклеотида. Это и есть сахаро-фосфатный скелет, от которого ответвляются отдельные пурины и пиримидины. Далее Левин показал, что сахара нуклеиновых кислот могут быть двух типов: рибоза, содержащая только пять атомов углерода вместо шести, как это имеет место в хорошо изученных сахарах, и дезоксирибоза, в которой на один атом кислорода меньше, чем в рибозе. Каждая молекула нуклеиновой кислоты содержит тот или иной сахар, но отнюдь не оба одновременно. Таким образом, различаются два типа нуклеиновых кислот: рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК). Каждая нуклеиновая кислота включает пурины и пиримидины четырех различных типов. В ДНК нет урацила, в ее состав входят А, Г, Ц и Т, в то время как в РНК нет тимина, а только А, Г, Ц и У. Шотландский химик Александр Тодд (род. в 1907 г.) подтвердил данные Левина, синтезировав в 40-х годах различные нуклеотиды. Вначале биохимики не придали большого значения нуклеиновым кислотам. Хотя и было известно, что белковая молекула связана с различными небелковыми дополнениями, вроде сахаров, жиров, металл- и витаминсодержащих соединений и т. д., считалось, что белок представляет собой основную часть молекулы. Даже после того, как нуклеопротеиды обнаружили в хромосомах и вирусах, биохимики не потеряли уверенности, что нуклеиновые кислоты — это второстепенная часть молекулы. В 90-х годах прошлого столетия Коссель провел наблюдение, все значение которого стало понятно гораздо позже. Сперматозоиды почти целиком состоят из тесно лежащих хромосом и содержат химические вещества, несущие полную информацию, благодаря которой потомству передаются отцовские наследственные признаки. Однако Коссель нашел, что белки сперматозоидов значительно проще, чем белки других тканей, в то время как нуклеиновая кислота подобна нуклеиновой кислоте тканей тела. Отсюда с большой вероятностью вытекало, что наследственная информация заключена скорее в неизменных молекулах нуклеиновых кислот спермы, чем в ее чрезвычайно упрощенном белке. Но вера в молекулу белка еще не была поколеблена, так как результаты исследований 30-х годов говорили о слишком простом, чтобы нести наследственную информацию, строении нуклеиновых кислот, представляющих очень мелкие молекулы, которые состоят только из четырех нуклеотидов. Поворотным пунктом явились исследования, проведенные в 1944 г. рядом ученых под руководством американского бактериолога Освальда Теодора Эвери (1877–1955), работавших со штаммами пневмококков (возбудителей пневмонии). У части штаммов была гладкая форма (S-штаммы) и наружная оболочка вокруг клетки (капсула), у другой — шероховатая без оболочки (R-штаммы). По-видимому, у R-штаммов отсутствовала способность синтезировать вещество капсулы. Вытяжка из S-штаммов, добавленная к R-штаммам, превращала последние в S-штаммы. Сама по себе вытяжка не может образовывать капсулы, но, по-видимому, вызывает такие изменения в R-штаммах, которые позволяют бактерии справиться с этой задачей. Вытяжка несет генетическую информацию, необходимую для изменения физических свойств бактерии. Но самая поразительная часть опыта выявилась при анализе вытяжки, которая представляла собой раствор, состоящий исключительно из нуклеиновой кислоты без примеси каких-либо белков. Итак, по крайней мере в этом случае, нуклеиновые кислоты, а не белок были генетическим материалом. С этого момента стало ясно, что именно нуклеиновая кислота — первичная и ключевая основа жизни. А так как в том же, 1944 г, впервые осуществили метод хроматографии на бумаге, то 1944 г., так же как и 1859 г., когда вышло в свет «Происхождение видов», можно справедливо назвать годом величайших биологических событий. Начиная с 1944 г. новый взгляд на нуклеиновые кислоты получил наибольшее обоснование благодаря исследованиям вирусологов. Было показано, что вирусы имеют внешнюю белковую оболочку, внутри которой находится молекула нуклеиновой кислоты. В 1955 г. американскому биохимику Гейнцу Френкель-Конрату (род. в 1910 г.) удалось разделить вирус на две составные части и вновь соединить их. Белковая часть сама по себе не обладала никакими инфекционными свойствами, она была мертва. Часть, содержащая нуклеиновую кислоту, была живой, инфекционной, хотя наибольшую активность проявляла в присутствии белкового компонента. Использование радиоактивных изотопов показало, что при внедрении бактериофага в бактериальную клетку проникает только та его часть, которая состоит из нуклеиновой кислоты, а белковая остается снаружи. Внутри клетки нуклеиновая кислота вызывает образование не только новых молекул нуклеиновой кислоты, подобных себе (а не нуклеиновой кислоте клетки), но и молекул белка, характерного для бактериофага, а не для клетки. Не остается никаких сомнений, что именно молекула нуклеиновой кислоты, а не белок несет генетическую информацию. Молекулы вируса содержат либо ДНК, либо РНК, либо обе нуклеиновые кислоты одновременно. В клетке, однако, ДНК обнаружена исключительно в генах. А поскольку гены являются единицами наследственности, окончательно проясняется значение ДНК. Структура нуклеиновой кислотыС работ Эвери началось энергичное изучение нуклеиновых кислот. Впечатление, что нуклеиновые кислоты представляют собой мелкие молекулы, создавалось потому, что ранние методы их выделения были достаточно грубы и расщепляли молекулу на более мелкие фрагменты. Более тонкие методы показали, что молекулы нуклеиновой кислоты крупнее даже самых больших молекул белка. Американский биохимик Эрвин Чаргафф (род. в 1905 г.) расщепил молекулу нуклеиновой кислоты и подверг ее фрагменты хроматографическому разделению на бумаге. В конце 40-х годов он показал, что в молекуле ДНК число пуриновых групп равно числу пиримидиновых. А если говорить конкретно, то число групп аденина (пурин) обычно равно числу тиминовых (пиримидин), в то время как количество групп гуанина (пурин) равно числу цитозиновых (пиримидин). Это можно выразить так: А-Т и Г-Ц. 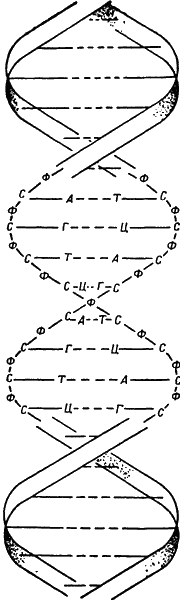 Рис. 7. Двойная спираль молекулы ДНК. Тяжи скелета состоят из чередующихся групп сахаров (С) и фосфатов (Ф). Внутрь ответвляются азотистые основания: аденин (А), гуанин (Г), тимин (Т) и цитозин (Ц). Пунктирные линии — водородные связи, которые соединяют тяжи спиралей. В процессе репликации каждый из скелетов воспроизводит свой комплект из пуринов и пиримидинов (А, Г, Ц, Т), которые всегда присутствуют в клетке. Английский физик Маурис Хью Фредерик Уилкинс (род. в 1916 г.) в начале 50-х годов применил к изучению ДНК метод дифракции рентгеновских лучей. Его коллеги по Кембриджскому университету английский биохимик Фрэнсис Гарри Комптон Крик (род. в 1916 г.) и американский биохимик Джеймс Дьюк Уотсон (род. в 1928 г.) попытались представить структуру молекулы так, чтобы она обьясняла данные Уилкинса. Незадолго до этого Полинг выдвинул гипотезу винтообразного спиралевидного строения белков. Крику и Уотсону казалось, что молекула ДНК, построенная по винтообразной спирали, соответствовала бы данным Уилкинса. Однако им пришлось допустить существование двойной винтообразной спирали, чтобы объяснить также и данные Чаргаффа. Образно они представляли себе молекулу состоящей из двух сахаро-фосфатных скелетов, извивающихся вокруг общей оси и создающих цилиндрическую форму молекулы. Внутрь, к центру цилиндра, ответвляются пурины и пиримидины. Для сохранения одинакового диаметра на всем протяжении цилиндра крупные молекулы пуринов должны примыкать к мелким молекулам пиримидинов, то есть А к Т, Г к Ц, что и объясняет данные Чаргаффа. Более того, возникает приемлемое объяснение основных стадий митоза, удвоения хромосом и близкого к этой проблеме механизма репродуцирования вируса в клетке. Каждая молекула ДНК образует отпечаток самой себя (реплику) следующим образом: оба сахаро-фосфатных скелета развертываются, и каждый служит моделью для нового комплекта. В каком бы месте скелета ни находился аденин, из наличных запасов клетки выбирается именно молекула тимина, и наоборот. Где бы ни размещалась молекула гуанина, она выбирает именно молекулу цитозина, и наоборот. Таким образом, скелет 1 образует новый скелет 2, в то время как скелет 2 образует новый скелет 1. Довольно быстро возникают две двойные винтообразные спирали там, где до этого существовала лишь одна. Если молекула ДНК реплицирует себя по всей длине хромосомы (или вируса), то процесс кончается образованием двух идентичных хромосом там, где была только одна. Но не всегда этот процесс протекает до конца вполне гладко. Если какое-либо воздействие нарушает ход репликации, новая молекула ДНК несколько отличается от своего «предка»; в этом случае мы имеем дело с мутацией. Модель молекулы ДНК, созданная Уотсоном и Криком, увидела свет в 1953 г. Генетический кодНо как молекула нуклеиновой кислоты передает информацию, касающуюся физических свойств? Ответ на этот вопрос дали результаты исследований американских генетиков Джорджа Уэлса Бидла (род. в 1903 г.) и Эдварда Лаури Тэтума (род. в 1909 г.). В 1941 г. они начали серию экспериментов с плесневым грибком Neurospora crassa, способным благодаря синтезу аминокислот из более простых азотсодержащих соединений жить на питательной среде, лишенной аминокислот. Если подействовать на плесень рентгеновскими лучами, возникают мутанты. Некоторые из них теряют способность продуцировать нужные аминокислоты. Один мутантный штамм, например, потерял способность образовывать аминокислоту лизин, и, чтобы поддержать жизнь этого штамма, ее приходилось вводить в питательную среду. Подобная дефективность, как показали Бидл и Тэтум, зависит от отсутствия специфического фермента, имеющегося у нормального немутантного штамма. Отсюда они сделали вывод, что способность продуцировать лизин представляет специфическую функцию особого гена, управляющего образованием данного фермента. Молекулы нуклеиновой кислоты, передаваемые через сперматозоид или яйцеклетку, обладают способностью продуцировать особый набор ферментов. Назначение этих ферментов — управлять химизмом клетки. Химизм клетки в свою очередь ответствен за все свойства, наследственность которых и изучали Бидл и Тэтум. Таким образом, можно было перекинуть мостик от ДНК к физическим признакам организма. Так как ДНК генов остается в пределах ядра, а синтез белка протекает вне ядра, образование ферментов генами, вероятно, проходит через промежуточные продукты. Электронно-микроскопическое изучение клетки раскрыло более тонкие детали ее строения и определило точное место белкового синтеза. В клетке в большом количестве были найдены организованные гранулы, значительно более мелкие, чем митохондрии, и потому названные микросомами (от греческих слов mikros — малый и soma — тело). В 1956 г. одному из наиболее энергичных исследователей микросом, американцу Джорджу Эмилю Паладе (род. в 1912 г.), удалось показать, что они богаты РНК (поэтому их переименовали в рибосомы). Тогда и обнаружили, что именно рибосомы являются местом синтеза белка. Но генетическая информация от хромосом должна дойти до рибосом. Это осуществляет особая разновидность РНК, названная информационной; информационная РНК точно повторяет структуру определенного участка ДНК хромосом, составляющего единицу наследственной информации — ген, и переносится из ядра в цитоплазму клетки, где и прикрепляется к рибосоме. Но для того, чтобы синтезировались белки, необходимы аминокислоты, которые образуются при помощи ферментов в самой клетке или поступают с пищевыми продуктами. Проблему доставки аминокислот в рибосомы впервые изучил американский биохимик Мелон Буш Хогленд (род. в 1921 г.). Он установил, что каждая аминокислота, прежде чем попасть к месту синтеза белков, соединяется с транспортной РНК, которая и переносит их на соответствующее место информационной РНК. Оставалось неясным: как молекула данной транспортной РНК прикрепляется к данной аминокислоте? Проще всего было бы представить, что аминокислота прикрепляется к пуринам и пиримидинам нуклеиновой кислоты; к каждому пурину или пиримидину — разные аминокислоты. Однако из 20 различных аминокислот молекулы белка на молекулу нуклеиновой кислоты приходится лишь четыре пурина и пиримидина. По этой причине вполне очевидно, что к каждой аминокислоте должна подходить комбинация по крайней мере из трех нуклеотидов. (Из трех нуклеотидов возможны 64 различные комбинации.) Подгон комбинаций тринуклеотидов к аминокислоте (то есть какая комбинация нуклеотидов и в какой последовательности в составе информационной РНК соответствует определенной аминокислоте) представлял самую важную биологическую проблему начала 60-х годов, относящуюся к расшифровке генетического кода. В этом направлении наиболее активно работает американский биохимик Северо Очоа (род. в 1905 г.). Происхождение жизниИтак, достижения молекулярной биологии к середине XX в. чрезвычайно сильно укрепили материалистические позиции. Всю генетику можно было истолковать с точки зрения химии, согласно законам, одинаково справедливым для живой и неживой природы. Даже мозг подвергался этому натиску. Вполне возможно, что процессы обучения и запоминания являются не только процессами возникновения и закрепления нервных путей, но и представляют собой синтез и сохранение специфических молекул РНК. Оставался незатронутым лишь один аспект биологии XIX в., в котором еще господствовала виталистическая точка зрения, — факт недоказанности самопроизвольного зарождения. Если формы жизни действительно никогда не могли развиться из неживой материи, как тогда возникла жизнь? Легче всего было предположить, что жизнь создана сверхъестественными силами. В 1908 г. шведский химик Сванте Август Аррениус (1859–1927) выдвинул гипотезу происхождения жизни без участия сверхъестественных сил. Он высказал мысль, что жизнь на Земле началась тогда, когда на нашу планету из космоса попали зародыши жизни. «Частицы жизни», носящиеся в бескрайних космических пространствах, переносимые давлением света от звезд, оседали то здесь, то там, осеменяя ту или иную планету. Гипотеза Аррениуса лишь отодвигала решение проблемы. Если жизнь была занесена на нашу планету извне, как она возникла там, откуда к нам попала? А может быть, жизнь все-таки возникла из неживой материи? Колбы Пастера сохранялись стерильными в течение какого-то ограниченного времени; а если их оставить на миллиарды лет? Или вместо колб представить целый океан раствора в условиях, далеких от современных? Нет причин думать, что основные химические вещества, складывающие живое, существенно менялись на протяжении веков. Весьма вероятно, что они не изменились. Действительно, аминокислоты, выделенные в небольших количествах из некоторых ископаемых организмов, насчитывающих десятки миллионов лет, оказались идентичными аминокислотам, встречающимся в живых тканях современных организмов. И все же химизм мира в целом мог измениться. Новые данные по химии Вселенной позволили американскому химику Гарольду Клейтону Ури (род. в 1893 г.) предположить, что первичная атмосфера Земли состояла из водорода и водородсодержащих газов, таких, как метан и аммиак; в ней совершенно отсутствовал свободный кислород, а значит, в ее верхних слоях не было озона (одной из форм кислорода). Сейчас такой слой озона существует и поглощает значительную часть ультрафиолетовых лучей солнечного света. В бедной первичной атмосфере несущая энергию радиация, возможно, проникала до океана, где и вызывала такие реакции, которых в настоящее время уже не может быть. Постепенно могли создаваться комплексы молекул; при отсутствии жизни они не потреблялись, а скапливались. В итоге реплицирующиеся молекулы создавали комплекс нуклеиновых кислот, и это было основой жизни. Благодаря мутациям и действию естественного отбора образовывались все более активные формы нуклеиновых кислот. Эти кислоты могли превратиться в клетки; последние, возможно, начали синтезировать хлорофилл. Фотосинтез (с помощью других процессов, в которые не вовлекались, вероятно, живые организмы) мог обогатить первичную атмосферу Земли свободным кислородом. А в такой атмосфере и в мире, где кишит жизнь, самопроизвольное зарождение описанного выше типа, вероятно, было бы уже невозможно. Эта гипотеза, хотя и тщательно продуманная, в значительной степени остается гипотезой. Однако в 1953 г. один из учеников Ури, Стенли Ллойд Миллер (род. в 1930 г.), поставил очень интересный опыт. Он взял тщательно очищенную и стерилизованную воду и добавил к ней «атмосферу» из водорода, аммиака и метана. Миллер заставлял эту смесь циркулировать в герметически изолированном приборе, через который пропускал электрические разряды, имитирующие ультрафиолетовое солнечное излучение. Опыт шел в течение недели, после чего Миллер разделил содержимое прибора методом хроматографии на бумаге. В растворе обнаружились простые органические соединения и даже несколько простейших аминокислот. В 1962 г. схожие опыты были повторены в Калифорнийском университете. К атмосфере добавляли этан (соединение, очень сходное с метаном, но содержащее два атома углерода). В результате было получено еще большее разнообразие органических соединений. И наконец, в 1963 г. подобным же образом синтезировали аденозинтрифосфат, один из основных высокоэнергетических фосфатов. Если это воспроизводимо в течение недели в небольшом приборе, чего же можно ожидать за миллиарды лет в огромном океане и одевающей его атмосфере? Очень трудно представить процесс эволюции на ранних этапах существования Земли, но когда мы попадем на Луну, мы, возможно, познакомимся с химическими процессами дожизненных формаций. А если доберемся до Марса, вероятно, изучим простые формы жизни, развивающиеся в условиях, совершенно отличных от условий Земли. Все это, несомненно, послужит углубленному пониманию земных проблем. Даже на нашей собственной планете мы с каждым годом узнаем все больше нового. В 1960 г. человек проник в самые недоступные глубины океана, в условия, совершенно чуждые ему. Вероятно, именно в океане мы установим контакт с нечеловеческим разумом, каким, по-видимому, обладают дельфины. А человеческий мозг — не раскроет ли он свои секреты, если подойти к его изучению с позиций молекулярной биологии? Благодаря успехам кибернетики и электроники, возможно, удастся создать неодушевленные мыслящие системы. Но зачем предаваться гаданиям, когда нужно только подождать? Пожалуй, в том и заключается великолепие науки, что в будущем она обещает гораздо более удивительное и великое, чем все, что сделано ею в прошлом. Сколько нового еще будет открыто нашими современниками? Рекомендуемая литература1. Архангельский Г. В., История неврологии от истоков до XX века, изд-во «Медицина», М., 1965. 2. Белозерский А. Н. и Микулинский С. Р., Успехи советской биологии, изд-во «Знание», М., 1967. 3. Бирюков Д. А. и Михайлов В. П., Эволюционно-морфологические и физиологические основы развития медицины за советский период, Медгиз, Л., 1957. 4. Завадовский М. М., Основные этапы истории экспериментальной биологии (зоологии) в России, Уч. зап. МГУ, вып. 103, т. 2, кн. I, М., 1946. 5. Из истории отечественной биологии XVIII–XIX веков, Сб. статей под ред. Коштоянца X. С. и Соболя С. Л., Изд-во АН СССР, М., 1953. 6. История естествознания в России, тт. 1–3, Изд-во АН СССР, М., 1957–1962. 7. История медицины СССР, Сб. статей под ред. Петрова Б. Д., изд-во «Медицина», М., 1964. 8. Карасик В. М., Прошлое и настоящее фармакологии и лекарственной терапии (исторический очерк), изд-во «Медицина», Л., 1965. 9. Коштоянц X. С., Очерки по истории физиологии в России, Изд-во АН СССР, М. — Л., 1946. 10. Лункевич В. В., От Гераклита до Дарвина. Очерки по истории биологии, изд. 2, тт. 1–2, М. — Л., 1960. 11. Люди русской науки (биология, медицина), Госиздат, Физматлит, М., 1963. 12. Майоров Ф. П., История учения об условных рефлексах, Изд-во АМН СССР, М. — Л., 1954. 13. Матвеев Б. С., Русская школа морфологов и ее роль в развитии дарвинизма, Уч. зап. МГУ, вып. 103, т. 2, кн. I, М., 1946. 14. Развитие биологии в СССР за 50 лет, изд-во «Наука», М., 1967. 15. Райков Б. Е., Очерки по истории эволюционной идеи в России до Дарвина, тт. 1–3, Изд-во АН СССР, М. — Л. 1947–1955. 16. Ручковский Б. С., Очерки развития советской экспериментальной онкологии, Изд-во АН УССР, К., 1959. 17. Старосельская-Никитина О. А., История естествознания. Литература, опубликованная в СССР, тт. 1–4, М. 1949–1955. 18. Толкачевская Н. Ф., Развитие биохимии животных, Краткий исторический очерк, Изд-во АН СССР, М., 1963. Оглавление Предисловие к русскому изданию… 5 Глава I. Древняя биология Зарождение науки… 7 Ионийская школа… 9 Афинская школа… 11 Александрийцы… 13 Эпоха римского владычества… 15 Глава II. Биология в средние века Мрачное время… 18 Эпоха Возрождения… 20 Переходный период… 23 Глава III. Рождение современной биологии Новая анатомия… 25 Кровообращение… 27 Начала биохимии… 31 Появление микроскопа… 32 Глава IV. Классификация живых форм Самопроизвольное зарождение… 36 Расположение видов в системе… 39 На подступах к теории эволюции… 41 Геологические предпосылки… 44 Глава V. Химия клетки Газы и жизнь… 48 Органические соединения… 51 Ткани и эмбрионы… 55 Глава VI. Эволюция Естественный отбор… 61 Борьба вокруг эволюционной теории… 65 Происхождение человека… 67 Боковые ветви эволюционной теории… 69 Глава VII. У истоков генетики Слабое место в теории Дарвина… 72 Горох Менделя… 73 Мутации… 77 Хромосомы… 79 Глава VIII. Конец витализма Азот и диета… 84 Калориметрия… 86 Брожение… 88 Ферменты… 91 Глава IX. Борьба с болезнями Вакцинация… 95 Микробная теория болезней… 97 Бактериология… 98 Насекомые… 101 Факторы питания… 103 Витамины… 106 Глава X. Нервная система Гипноз… 109 Нервы и головной мозг… 112 Поведение… 115 Нервные потенциалы… 118 Глава XI. Кровь Гормоны… 121 Серология… 124 Группы крови… 125 Вирусные заболевания… 128 Аллергия… 132 Глава XII. Обмен веществ Химиотерапия… 135 Антибиотики и пестициды… 137 Продукты промежуточного обмена… 140 Радиоактивные изотопы … 145 Глава XIII. Молекулярная биология: белок Ферменты и коферменты… 148 Электрофорез и дифракция рентгеновских лучей… 150 Метод хроматографии… 154 Расположение аминокислот… 156 Глава XIV. Молекулярная биология: нуклеиновая кислота Вирусы и гены… 159 Роль ДНК… 162 Структура нуклеиновой кислоты… 165 Генетический код… 168 Происхождение жизни… 170 Рекомендуемая литература… 173 |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Наверх |
||||
|
|
||||