|
||||
|
|
Глава VIIУ истоков генетики Слабое место в теории ДарвинаПроцесс передачи потомству родительских наследственных факторов долгое время оставался совершенно непонятным. Открытие в конце XVII в. сперматозоида вызвало целую дискуссию. Одни утверждали, что будущий зародыш целиком заключен в яйцеклетке и оплодотворение является лишь толчком к развитию. Другие настаивали на том, что будущий зародыш помещается в сперматозоиде, а яйцеклетка лишь обеспечивает его питание. Спорщики сходились на том, что носителем наследственных факторов является одна родительская особь. Только в середине XVIII в. в результате наблюдений над детьми от смешанных браков у человека и исследования экстерьера мулов было установлено, что признаки наследуются от обоих родителей. Пьер Луи Моро Мопертюи (1698–1759) выдвинул теорию, согласно которой наследственные признаки у потомков формируются и определяются «семенными частицами» обоих родителей. Даже в XIX в. еще не было правильного представления о наследственном механизме. Именно поэтому эволюционная теория так часто применялась неудачно. Спенсер считал возможным быстрое изменение человеческого поведения: Гальтону казалось весьма несложным усовершенствовать человеческую расу с помощью подбора производителей при размножении. Подобные воззрения биологов объяснялись, по сути дела, недостатком знаний о природе наследственного механизма. Это было самое слабое звено в теории Дарвина. Дарвин предполагал, что среди молодых особей любого вида наблюдается непрерывная случайная изменчивость, причем определенные изменения делают животных более приспособленными к окружающей среде. Так, жирафу тем легче прокормиться, чем длиннее у него шея. Но где гарантия, что этот признак повторится у потомства? Вряд ли жираф выискивал особо длинношеего супруга, гораздо вероятнее, что ему пришлось встретиться с короткошеим. При скрещивании крайних вариантов происходит смешение признаков, так что у потомства длинношеего и короткошеего жирафов шея будет средней длины, — к такому убеждению привел Дарвина весь его опыт по разведению животных. Иными словами, в результате случайного спаривания все полезные и приспособительные признаки, которые проявляются при случайной изменчивости, усредняются. Раз естественному отбору нечего «отбирать», то и эволюционных изменений происходить не будет. Попытки биологов объяснить это явление оказались безуспешными. Немецкий ботаник Карл Вильгельм Негели (1817–1891) прекрасно сознавал, как сложно найти четкое обоснование «усреднения» и его последствий. Он предположил, что эволюционные изменения направляются в определенную сторону каким-то внутренним толчком. Так, судя по палеонтологическим данным, предками лошадей были животные величиной с собаку, имевшие по четыре копытца на каждой стопе. С течением времени эти животные становились крупнее и утрачивали одно копытце за другим, пока не превратились в современную крупную однокопытную лошадь. Негели предполагал, что побудительным фактором этого процесса было действие некой внутренней силы. Если бы она действовала и дальше, лошади могли бы стать слишком крупными и неуклюжими и, следовательно, беззащитными против врагов; им грозило бы постепенное вымирание. Эта теория носит название теории ортогенеза. Современные биологи ее отвергают. Как мы увидим дальше, приверженность Негели к этой теории неожиданно принесла скверные плоды. Горох МенделяПроблема была решена благодаря трудам чешского натуралиста Грегора Иоганна Менделя (1822–1884). Мендель занимался и математикой и ботаникой. Начиная с 1856 г. он в течение девяти лет изучал наследственные признаки гороха, используя статистические приемы обработки результатов. Ученому нужна была полная уверенность, что наследуются признаки только одного родителя, поэтому он очень тщательно проводил самоопыление различных растений, аккуратно собирал с каждого самоопыленного растения семена, отдельно высаживал их и изучал новое поколение. В результате этих опытов Мендель обнаружил, что горошины от карликовых растений и в первом и в последующих поколениях дают только карликовые растения. Таким образом, карликовые растения чистосортны. Высокорослые растения вели себя по-разному. Некоторые (более трети) оказались чистосортными и поколение за поколением давали высокие растения. Часть остальных семян давала высокие, а часть — карликовые растения, причем высокорослых всегда получалось в два раза больше, чем карликовых. Очевидно, имелось два типа высоких растений — чистосортные и нечистосортные. Мендель пошел дальше. Он скрестил карликовые растения с чистосортными высокорослыми и обнаружил, что из каждого гибридного семени вырастает высокое растение. Казалось бы, признаки карликовости исчезли. Произведя затем самоопыление каждого гибридного растения, Мендель изучил полученные семена. Все гибридные растения оказались нечистосортными. Около одной четверти горошин дали карликовые растения, одна четверть — чистосортные высокорослые, а оставшаяся половина — нечистосортные высокорослые растения. Мендель предположил, что каждое растение гороха несет два фактора, определяющих какой-либо признак, в данном случае — высоту. Один фактор содержится в пыльце, другой — в семяпочках. После оплодотворения новое поколение содержит уже оба фактора (по одному от каждого родителя, если произошло скрещивание двух растений). У карликовых растений имеются только факторы «карликовости»; при комбинации их путем перекрестного опыления или самоопыления получаются только карликовые растения. Чистосортные высокорослые растения содержат только факторы «высокорослости», и их комбинации дают только высокие растения. 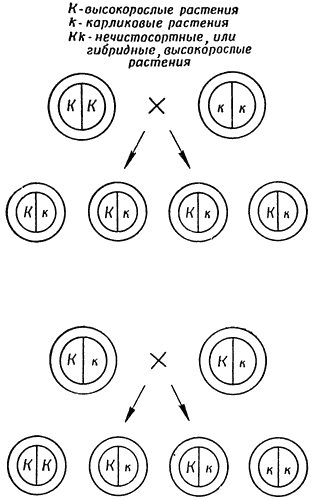 Рис. 3. Схема, объясняющая опыты Менделя по скрещиванию высокорослых и карликовых растений гороха. Вверху — скрещивание чистосортного высокорослого растения с карликовым, которое дает гибридные, или нечистосортные, высокорослые растения. Внизу — скрещивание между собой гибридных растений первого поколения, которое дает чистосортные высокорослые растения, гибридные высокорослые растения и карликовые растения в отношении 1:2:1. Фактор карликовости обозначается буквой к, так как он рецессивен, а соответственный ему доминантный фактор высокорослости — К. Когда чистосортные высокие растения скрещиваются с карликовыми, факторы «высокорослости» и «карликовости» комбинируются и следующее поколение оказывается гибридным. Все гибриды будут высокими, так как фактор «высокорослости» является доминантным, маскирующим действие фактора «карликовости». Последний, однако, не исчезает, а сохраняется. Такие гибриды не будут вести себя как чистосортные, потому что содержат оба фактора, комбинации которых определяет чистая случайность. Фактор «высокорослости» может комбинироваться с другим фактором «высокорослости», тогда появится чистосортное высокорослое растение. Так произойдет в одной четверти случаев. В другой четверти фактор «карликовости» в комбинации с другим фактором «карликовости» дает карликовое растение. В оставшейся половине фактор «высокорослости» сочетается с фактором «карликовости» или фактор «карликовости» — с фактором «высокорослости»; при этом получаются высокорослые нечистосортные растения. Мендель показал, что наследование любого признака можно объяснить таким же образом, причем для всех изученных им признаков скрещивание двух крайностей не приводило к смешению наследственности — каждый вариант сохранялся без изменения; если он исчезал в одном поколении, то появлялся в следующем. Все это имело немаловажное значение для эволюционной теории (хотя сам Мендель никогда не думал о приложении к ней своих идей), так как позволяло сделать следующий вывод. Случайные вариации, в течение какого-то промежутка времени возникающие внутри вида, в конечном счете не усредняются, а исчезают и проявляются вновь, пока их в полной мере не использует естественный отбор. Причина, по которой унаследованные признаки часто выглядят промежуточными, состоит в том, что при скрещивании растений и животных большинство признаков обычно является комбинацией нескольких факторов. Различные компоненты наследуются независимо, но в то же время по принципу «да» или «нет». Среднее количество «да» или «нет» определяет появление промежуточного признака. Открытия Менделя касались и евгеники. Искоренять нежелательные признаки оказалось не так легко, как представлялось вначале. Не проявившись в одном поколении, они могут обнаружиться в следующем. Размножение, сопровождаемое отбором, — процесс более тонкий и более продолжительный, чем думал Гальтон. Однако мир был еще не подготовлен к тому, чтобы оценить эти факты. Тщательно описав результаты своих экспериментов, Мендель решил заинтересовать какого-нибудь известного ботаника, чтобы заручиться его поддержкой. Поэтому в начале 60-х годов XIX в. он переслал свои работы Негели. Тот прочел рукопись и отозвался весьма холодно. Теория, основанная на подсчетах растений гороха, не произвела на него никакого впечатления. Он предпочитал туманный и многословный мистицизм, который так характерен для его собственного учения об ортогенезе. Неудача обескуражила Менделя. В 1866 г. он опубликовал свою статью, но исследований уже не продолжал. Да и статья не обратила на себя особого внимания, поскольку Негели не поддержал Менделя. Менделя с полным правом можно назвать основателем учения о механизмах наследственности, которое теперь называется генетикой, но в то время никто этого не предполагал, в том числе и он сам. МутацииВо второй половине XIX столетия возникла другая проблема, связанная с развитием эволюционных идей. В результате новейших открытий, сделанных физиками, появилось неожиданное представление о значительно меньшей длительности истории Земли. С провозглашением закона сохранения энергии возник вопрос об источнике солнечной энергии. О радиоактивности и ядерной энергии стало известно много позже, так что в течение XIX в. не было дано удовлетворительных доказательств того, что Солнце пребывало в его теперешнем состоянии на протяжении по крайней мере десятков миллионов лет. Для эволюции по дарвиновскому типу времени было явно недостаточно, и некоторые биологи, в том числе Негели и Келликер, предположили, что эволюция идет скачками. Хотя потом выяснилось, что масштаб времени определен неправильно, мысль о скачкообразности эволюционного процесса все же оказалась плодотворной. Голландский ботаник Гуго де Фрис (1848–1935) также принадлежал к тем, кто заговорил о скачках в эволюции. Как-то на заброшенном лугу ему попалась на глаза заросль ослинника — растения, незадолго до этого завезенного в Нидерланды из Америки. Наблюдательный глаз ботаника отметил, что некоторые из этих растений резко отличались по внешнему виду, хотя и происходили, вероятно, от общего предка. Он перенес ослинник в сад, стал разводить каждую форму отдельно и постепенно пришел к выводам, которые за много лет до него сделал Мендель: индивидуальные свойства передаются из поколения в поколение, не претерпевая смешения и усреднения; время от времени появляются новые признаки, заметно отличающие эти растения от других и сохраняющиеся в следующих поколениях. Де Фрис назвал такие внезапные изменения мутациями (от латинского mutatio — изменение) и решил, что он наблюдает скачкообразно протекающую эволюцию вида. В действительности мутации ослинника не были связаны с изменениями самих наследственных факторов. Вскоре, однако, были изучены и настоящие мутации. Подобные явления уже давно были известны пастухам и фермерам, часто наблюдавшим появление уродств и всякого рода отклонений от нормального типа. Некоторые такие отклонения даже использовались в сельскохозяйственной практике. Например, мутация «коротконогости» овец появилась в Новой Англии в 1791 г. Короткие ноги не позволяли овцам перепрыгивать даже через очень низкую изгородь. Этих овец начали усиленно разводить и оберегать. Но пастухи, как правило, не делают теоретических выводов из своих наблюдений, а ученые, к сожалению, часто незнакомы с практическим скотоводством. И только благодаря де Фрису эти явления наконец предстали перед научным миром. Около 1900 г., готовясь к опубликованию своих открытий и просматривая старые работы, посвященные этой проблеме, ученый, к своему удивлению, обнаружил статью Менделя тридцатилетней давности. Независимо от де Фриса еще два ботаника — немец Карл Эрих Корренс (1864–1933) и австриец Эрих Чермак (род. в 1871 г.) — в один и тот же год пришли к весьма сходным выводам. И каждый из них, просматривая предшествующие работы, обнаружил статью Менделя. Все трое, де Фрис, Корренс и Чермак, опубликовали свои работы в 1900 г., и каждый, ссылаясь на работу Менделя, оценивал свои исследования как простое подтверждение его выводов. Поэтому мы и говорим теперь о менделевских законах наследственности. Эти законы в сочетании с открытием де Фриса дают картину возникновения и сохранения вариаций. Так было устранено слабое место в теории Дарвина. А после того, как английский ученый Рональд Фишер (1890–1962) в своей книге «Генетическая теория естественного отбора» (1930) показал, что дарвиновская теория отбора и менделевская генетика должны быть сведены в единую теорию эволюции, были достигнуты заметные успехи. В позднейших работах англичанина Джулиана Хаксли (род. в 1887 г.) и американца Джорджа Гейлорда Симпсона (род. в 1902 г.) показано, что отбор — более важный фактор эволюционного процесса, чем мутации. ХромосомыК 1900 г. менделевские законы приобретают гораздо большее значение в связи с новыми важными открытиями в области изучения клеток. В XVIII и в начале XIX столетий немногое можно было разглядеть в клетке, даже пользуясь усовершенствованным микроскопом. Клетка представляет собой довольно прозрачное тело и при наблюдениях имела вид более или менее резко очерченной капли. Биологам приходилось довольствоваться описанием ее внешней формы и размеров. Правда, иногда удавалось разглядеть в центре клетки несколько более плотное тело (теперь его называют клеточным ядром). В 1831 г. шотландский ботаник Роберт Браун (1773–1858) впервые предположил, что оно обязательно для каждой клетки. Семью годами позднее Шлейден в своей клеточной теории отводит ядру клетки весьма серьезную роль. Он связывает его с клеточным размножением, считая, что новые клетки отпочковываются от поверхности ядра. К 1846 г. Негели удалось доказать, что это неверно. Однако интуиция лишь частично подвела Шлейдена: ядро действительно связано с размножением клетки. Но исследование роли ядра требовало новой техники, которая позволила бы заглянуть внутрь клетки. Такую технику принесла органическая химия. Вслед за Бертло химики-органики быстро научились изготовлять органические вещества, не существующие в природе, — многие из них обладали яркой окраской. С 50-х годов XIX в. начался бурный рост производства синтетических красителей. Если считать содержимое клетки неоднородным, можно предположить, что отдельные ее части будут по-разному реагировать с различными химическими веществами. В результате обработки красителями некоторые части клетки окрасятся, а другие останутся бесцветными. Невидимые до тех пор детали благодаря окраске станут доступными наблюдению. Можно рассказать о многих биологах, экспериментировавших в этом направлении, но наиболее выдающихся успехов достиг немецкий цитолог Вальтер Флемминг (1843–1905). Он исследовал тонкое строение клетки животных и с помощью разработанных им методов фиксации и окраски обнаружил, что частицы, разбросанные внутри клеточного ядра, представляют собой образования, которые хорошо абсорбируют краситель, отчетливо выделяясь на бесцветном фоне. Флемминг назвал содержимое этих окрашивающихся частиц хроматином (от греческого chröma — окраска). Обрабатывая срезы растущей ткани, Флемминг, конечно, убивал клетки, причем каждая из них фиксировалась на той или иной стадии процесса деления. В 70-х годах XIX в. Флеммингу удалось выявить картину изменений хроматинового материала на всех этапах клеточного деления. Он обнаружил, что в начале процесса хроматиновый материал сливается, формируя короткие нитевидные образования, которые впоследствии получили название хромосом (от греческих chroma — окраска, soma — тело). Поскольку эти нитевидные хромосомы были типичны для процесса деления клетки, Флемминг назвал весь процесс митозом (от греческого mitos — нить). В клетке имеется небольшое тельце, носящее название центриоль, — оно напоминает звезду с расходящимися во все стороны лучами. Центриоль делится на две части, которые расходятся к противоположным концам клетки. Тонкие лучи, идущие от одной центриоли к другой, как бы опутывают хромосомы, группирующиеся в плоскости, проходящей через центр клетки.  Рис. 4. Митоз в клетке гипотетического животного. А — стадия покоя; Б — ранняя профаза (центриоль разделилась, появляются хромосомы); В — более поздняя профаза (хромосомы раздвоились, но еще остаются связанными); Г — поздняя профаза (ядерная оболочка растворилась, имеется веретено); Д — метафаза (хромосомы расположились по экватору веретена); Е — анафаза (хромосомы движутся к полюсам); Ж — телофаза (образовались ядерные оболочки, хромосомы удлиняются, начинается деление цитоплазмы); З — дочерние клетки (стадия покоя). 1 — центриоль; 2 — ядрышко; 3 — хроматин. Критический момент клеточного деления наступает, когда хромосома воссоздает свою собственную копию. Сдвоенные хромосомы растягиваются в противоположные стороны: одна хромосома каждой пары — в один конец клетки, другая — в другой; после этого клетка делится — в центре образуется перегородка. Каждая из двух новых дочерних клеток благодаря удвоению хромосом содержит такое же количество хроматина, какое было в исходной материнской клетке. Эти наблюдения Флемминг опубликовал в 1882 г. В 1887 г. бельгийский цитолог Эдуард ван Бенеден (1846–1910) наглядно показал две важные особенности хромосом. Во-первых, число хромосом в различных клетках организма постоянно, то есть каждый вид характеризуется определенным хромосомным набором (например, каждая клетка человека имеет 46 хромосом). Во-вторых, при образовании половых клеток — яйцеклетки и сперматозоида — в одном из делений не происходит удвоения хромосом. Следовательно, каждое яйцо и сперматозоид получают только половину типичного для вида количества хромосом. Второе рождение менделевского закона заставило по-новому взглянуть на исследования хромосом. В 1902 г. американский цитолог Уолтер Саттон (1876–1916) обратил внимание на то, что хромосомы ведут себя подобно менделевским наследственным «факторам»: каждая клетка имеет постоянное число пар хромосом. Они, видимо, несут в себе способность передавать физические признаки от клетки к клетке, так как в каждом клеточном делении число хромосом строго сохраняется; каждая хромосома создает копию (реплику) самой себя для использования ее в новой клетке. В оплодотворенной яйцеклетке, образовавшейся от слияния яйца и сперматозоида, восстанавливается прежнее число хромосом. При прохождении последовательных стадий деления в оплодотворенной яйцеклетке число хромосом опять строго сохраняется вплоть до образования самостоятельно живущего организма. Однако не следует забывать, что в новом организме одна хромосома из каждой пары получена от матери (через яйцеклетку), а другая — от отца (через сперматозоид). Это перемешивание хромосом, происходящее в каждом поколении, может вывести на свет рецессивные признаки, ранее подавленные доминантными. Новые комбинации в дальнейшем создают все новые вариации признаков, которые и «подхватываются» естественным отбором. Казалось, на заре XX столетия наступил небывалый расцвет эволюционного учения и генетики, но это было лишь прелюдией к новым, еще более поразительным достижениям. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Наверх |
||||
|
|
||||